




Hóa Chất Himitech

Hóa CHất Ngành Dược

HÓA CHẤT NGÀNH CÔNG NGHIỆP

Dung Môi Công Nghiệp

Hóa Chất Ngành Khai Khoáng

Hóa Chất Ngành Gốm Sứ, Thủy Tinh

HÓA CHẤT NGÀNH CÔNG NGHIỆP

HÓA CHẤT NGÀNH CÔNG NGHIỆP

Hóa Chất Ngành Dệt Nhuộm

Hóa Chất Ngành Thực Phẩm

HÓA CHẤT NGÀNH CÔNG NGHIỆP

HÓA CHẤT NGÀNH CÔNG NGHIỆP

HÓA CHẤT NGÀNH CÔNG NGHIỆP

Dung Môi Công Nghiệp

Hóa Chất Ngành Thực Phẩm

HÓA CHẤT NGÀNH CÔNG NGHIỆP

HÓA CHẤT NGÀNH CÔNG NGHIỆP

HÓA CHẤT NGÀNH THỰC PHẨM, DƯỢC, MỸ PHẨM

HÓA CHẤT NGÀNH THỰC PHẨM, DƯỢC, MỸ PHẨM

HÓA CHẤT NGÀNH CÔNG NGHIỆP

HÓA CHẤT NGÀNH CÔNG NGHIỆP

HÓA CHẤT NGÀNH CÔNG NGHIỆP

HÓA CHẤT NGÀNH CÔNG NGHIỆP
Hóa Chất Ngành Thực Phẩm

HÓA CHẤT NGÀNH CÔNG NGHIỆP

Hóa Chất Hãng Duksan

Thiết Bị Phòng Thí Nghiệm

Hóa Chất Hãng Acros Organics

Dụng Cụ Phòng Thí Nghiệm

Hóa Chất Hãng Thermo Fisher

Hóa Chất Hãng Thermo Fisher

Hóa Chất Hãng China

Hóa Chất Hãng Acros Organics

Thu Gom Phế Liệu

Vận Chuyển Hóa Chất

Dụng Cụ Chứa Đựng Hóa Chất

Thu Gom Phế Liệu
- 1
- 2
TIN TỨC
‘Đường sắt cao tốc Bắc – Nam là dự án cống hiến’
Đó là khẳng định của bà Đào Thụy Vân – Phó tổng giám đốc Công [...]
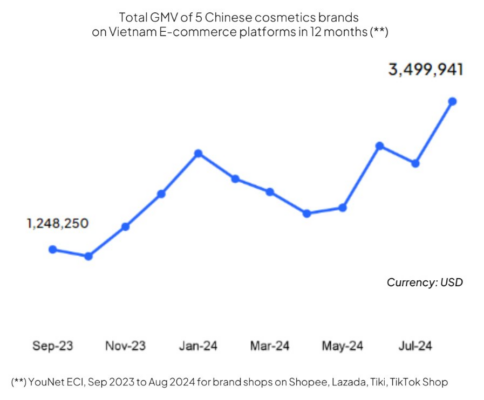
Mỹ phẩm nội địa Trung Quốc âm thầm “đổ bộ” Việt Nam: Chỉ 5 thương hiệu đã thu về hơn 660 tỷ đồng/năm nhờ một thế hệ “dễ mở ví, nhanh thay lòng”
Dữ liệu từ 4 sàn thương mại điện tử cho thấy Top 5 thương hiệu [...]
14
Th12
Th12

Chuyên gia nêu 4 đề xuất quan trọng thúc đẩy phát triển toàn diện Khu thương mại tự do Đà Nẵng
Chuyên gia nêu 4 đề xuất quan trọng thúc đẩy phát triển toàn diện Khu [...]
14
Th12
Th12

Không phải Trung Nguyên của ông Đặng Lê Nguyên Vũ, “ông vua” xuất khẩu cà phê Việt Nam là ai?
Chi nhánh CTCP Tập đoàn Trung Nguyên - Nhà máy cà phê Sài Gòn đứng [...]
14
Th12
Th12

